Per una miglior comprensione invitiamo a leggere due degli articoli precedenti:
RIPRODUZIONE IN CEPPI DIVERSI
Per ceppo si intende un individuo che ha caratteristiche genetiche definite e con fenotipo che rientra nel range della specie. Diventa importante parlare di ceppo in quanto, in caso di organismi di interesse enologico come i lieviti, diversi ceppi denotano differenze significative in termini tecnologici.
C’è una differenza interessante (non dal punto di vista enologico)sulla polarità di alcuni ceppi: alcuni lieviti riescono a cambiare la polarità a seconda delle condizioni ambientali in cui sono posti mentre altri non riescono a cambiare la polarità mantenendola per tutta la fase vegetativa aploide. Questi ceppi si dicono rispettivamente CEPPI OMOTALLICI e CEPPI ETEROTALLICI.
| CEPPI OMOTALLICI
POSSONO CAMBIARE LA POLARITÀ DELLA CELLULA APLOIDE |
CEPPI ETEROTALLICI
MANTENGONO LA POLARITÀ DELLA CELLULA APLOIDE SENZA LA POSSIBILITÀ DI CAMBIARE |
Questa differenza si basa sulla differente stabilità delle spore in fase aploide in quanto i ceppi eterotallici possono essere mantenuti in fase aploide in modo stabile mentre i ceppi omotallici NON possono essere mantenuti in maniera stabile in fase aploide
CEPPI OMOTALLICI
Ma come fanno effettivamente questi ceppi a cambiare polarità?
Questo è possibile perché nel cromosoma 3 sono presenti 3 tratti di DNA che contengono delle informazioni essenziali per attuare questo processo.
Su uno stesso cromosoma 3 di un determinato lievito aploide omotallico troviamo tre sequenze che potenzialmente potrebbero esprimere fattori sessuali.
Le sequenze sono composte da due segmenti principali:
– X: sequenza che esprime un gene che lavora in meiosi;
– Y: Ya ed Yα che esprimono rispettivamente il FSa ed il FSα.
Più precisamente possiamo fare una divisione di lavoro tra le cellule MATa e MATα: nelle MATa abbiamo l’informazione del fattore sessuale portata da Ya che andrà a codificare per il precursore di FSa, a1, dove poi una proteinasi[1] specifica lo trasformerà nella forma attiva; X, in questo caso, codifica per a2, elemento in grado di bloccare i geni di α. Nelle MATα abbiamo il segmento Yα e quattro copie dello stesso gene sono tradotte come polipeptide PRE-PRO-α1-α1-α1-α1, dove il PRE servirà per l’arrivo all’ER e il PRO il trasferimento al Golgi per far maturare la catena polipeptidica e liberare i 4 peptidi attivi; X codificherà per α2, in grado di bloccare i geni a.
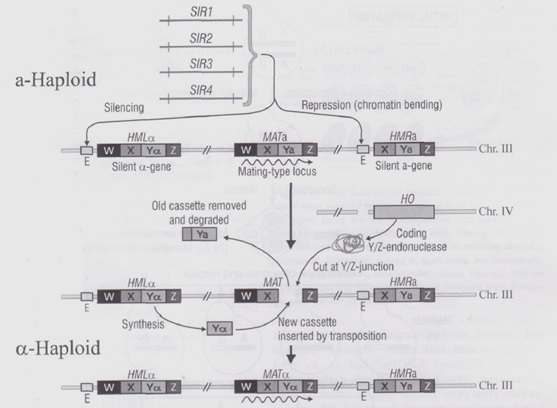
Se consideriamo un lievito MATa troveremo che al centro della zona del cromosoma 3 che interessa a noi possiamo notare una sequenza W – X – Ya – Z, codificata ed esprimente il FSa.
Le zone W e Z sono zone che delimitano le due importantissime sequenze X e Y , non codificano nulla e servono per localizzare le due sequen ze principali.
ze principali.
A valle di questa sequenza è riproposto il locus esprimente il FSa, chiamato locus HMRa, composto dalle sequenze X – Ya – Z; a monte, invece, è posto il locus HMLα, che, a differenza delle altre due sequenze, contiene la sequenza Yα, capace di esprimere il FSα, quindi la polarità opposta rispetto allo stato della cellula.
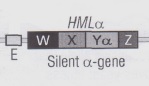
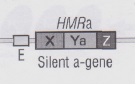
Le sequenze poste a monte e a valle del locus centrale MATa sono precedute da un pezzettino di DNA chiamato E, un target di un fattore di trascrizione che serve a bloccare la sintesi dell’RNA dei filamenti HMLα ed HMRa dipendentemente dalla sintesi dei geni SIR che legheranno il fattore E.
Da qua possiamo capire che i lieviti portano le informazioni sia per il FSa che per l’FSα visto che se una cellula si presenta come MATa sarà bloccato il FSα, mentre se una cellula si presenta come MATα sarà bloccato l’FSa; ma allora cos’è che permette ad un lievito di cambiare o meno la polarità?
Perché avvenga il cambio di polarità è necessario che la cellula esprima il fattore HO posto sul cromosoma 4 ed è proprio l’espressione di questo gene che permette il cambio di polarità e quindi la divisione tra lieviti omotallici ed eterotallici.
Il fattore HO codifica per un enzima chiamato YZendonuclease che taglia il DNA del cromosoma 3 in un punto particolare, ossia esattamente la sequenza Y, posta tra X e Z ed essenziale per il FS del lievito.
Una volta tagliata nel cromosoma si formerà un buco che attiverà un meccanismo di riparazione del DNA : la macchina di riparazione si attiverà, il cromosoma ripiegherà e si srotolerà tra la zona W e Z la quale verrà appaiata alle sequenza a monte nei loro rispettivi W e Z. Allineati i due tratti W e Z che sono posti uno dopo l’altro e quindi, in questo caso, appaiati, verrà utilizzata la sequenza Y presente sul locus HMLα (in questo caso) cambiando, di fatto, la sequenza Ya di prima in Yα.
È cosi avvenuto il cambio di polarità in quanto si esprimerà ora il FSα rendendo quella che prima era una cellula aploidea una cellula aploideα.
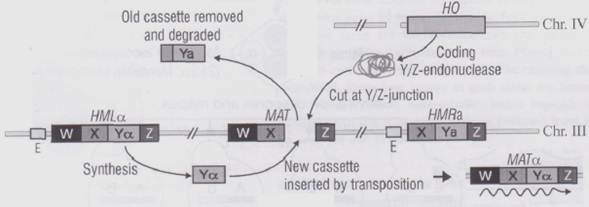
CEPPI ETEROTALLICI
I CEPPI ETEROTALLICI presentano una mutazione del gene Ho e, per questo, non possono effettuare cambiamenti di polarità. Da notare che nei lieviti l’eterotallismo è l’eccezione.
Ma perché è un vantaggio essere omotallici? Uno dei vantaggi maggiori è che una popolazione, senza riproduzione sessuale, potrebbe fissare, con la sola mitosi, caratteri recessivi che porterebbero alla morte della colonia (caso estremo) o comunque risulterebbe svantaggiata rispetto alle altre.
L’omotallismo serve ai lieviti come RINNOVAMENTO GENOMICO che avviene appunto grazie al cambiamento di polarità e alla riuscita della meiosi; infatti nel caso in cui venissero isolati ceppi con carattere recessivo, questi verrebbero eliminati (insieme al carattere negativo), mentre se il carattere sarà vantaggioso soppianterà i ceppi sprovvisti.
Le differenze tra meiosi e mitosi sono:
– MITOSI: si ha una prima duplicazione del DNA, dove è possibile che nella copiatura del DNA si introduca qualche mutazione, seguita dalla formazione di cromatina. Si forma, poi, un cromosoma a 4 bracci, il fuso mitotico e i filamenti di actina che verranno attaccati al centromero in modo da avere la separazione 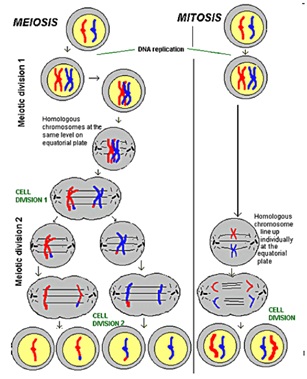 delle coppie dei singoli cromosomi e la citodieresi. Si conclude con la formazione di 2 cellule identiche (salvo mutazioni per errori durante la replicazione).
delle coppie dei singoli cromosomi e la citodieresi. Si conclude con la formazione di 2 cellule identiche (salvo mutazioni per errori durante la replicazione).
– MEIOSI: è fondamentale soprattutto per i processi di crossing over, più frequenti dove sono presenti determinate regioni chiamate hot-spot (soprattutto verso la parte più interna dei cromatidi mentre le parti esterne rimangono più simili ai parentali).
Si ha una prima duplicazione del DNA (meiosi I) e, subito dopo, il verificarsi del fondamentale processo di crossing over (hot spot). La formazione di cromatidi fratelli è poi seguita da una divisione senza sintesi di nuovo DNA (meiosi II) per finire con la formazione di 4 cellule aploidi (ascospore) le quali 2 saranno uguali ai parentali mentre 2 saranno diverse causa il fenomeno di crossing over.
Un esempio potrebbe essere la TRASLOCAZIONE di materiale genetico che attiva un gene responsabile della resistenza alla solforosa, l’ SSU1; per l’attivazione di questo gene c’è uno scambio tra cromosomi non omologhi 8 – 16 che fa si che questo gene venga controllato da un altro promotore.
Il gene verrà quindi espresso portano alla formazione di proteine di membrana che porteranno fuori dal lievito la solforosa (prodotta dal lievito).
Quando andiamo a selezionare i lieviti la variabilità genetica che è mantenuta dai processi di riproduzione sessuale è fondamentale per noi; infatti è solo grazie alla presenza di variabilità che si rende possibile la selezione di ceppi più o meno portati alla fermentazione e alla produzione di aromi.
Noi, per esempio, tramite incrocio, possiamo ottenere lieviti più vocati alla fermentazione e con spiccata produzione per aromi, mentre in natura possiamo trovare lieviti con o una o l’altra caratteristica.
Per i lieviti selezionati è fondamentale la STABILITÀ GENETICA, data soprattutto dalla forma diploide del lievito (autodiploidizzazione[1]); tutte le sorgenti di variabilità come crossing over, trasposoni o retrotrasposoni, traslocazioni ecc.. sono fonte di instabilità genetica.
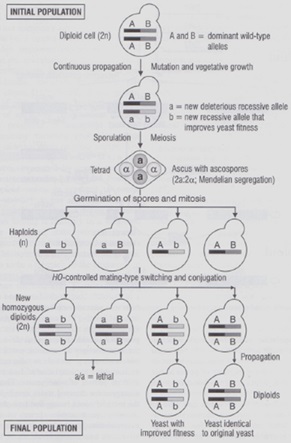 Infatti quando andiamo a coltivare un lievito in assenza di riproduzione sessuale provocheremo degli accumuli di mutazioni recessiva in forma eterozigote. Nei lieviti omotallici il “rinnovamento genomico” mediante il cambiamento della polarità porta alla formazione di omozigoti e se il carattere recessivo sarà danno saranno eliminati, mentre se il carattere sarà vantaggioso questi potranno soppiantare la popolazione di partenza.
Infatti quando andiamo a coltivare un lievito in assenza di riproduzione sessuale provocheremo degli accumuli di mutazioni recessiva in forma eterozigote. Nei lieviti omotallici il “rinnovamento genomico” mediante il cambiamento della polarità porta alla formazione di omozigoti e se il carattere recessivo sarà danno saranno eliminati, mentre se il carattere sarà vantaggioso questi potranno soppiantare la popolazione di partenza.
[1] Proteinasi : un enzima che sia in grado di catalizzare la rottura del legame peptidico tra il gruppo amminico e il gruppo carbossilico delle proteine
[1] AUTODIPLOIDIZZAZIONE: nei ceppi OMOTALLICI, dopo un certo tempo dalla germinazione delle spore, la spora stessa passa dalla forma aploide a quella diploide. Questo avviene grazie alla presenza di una sequenza HO codificata nel cromosoma 4 che permette il cambio di sesso della spora dandole la possibilità di combinarsi con un’altra presente nell’asco della cellula madre per creare un ibrido omozigote (diploide), dato che deriva dall’unione di due cellule aploidi uguali.
Buona lettura
Vino&Viticoltura